
Crossbreeding
- Breeding Systems A tool to Help Manage Simple Genetic Traits
- What is Crossbreeding?
- Improving Cow Herd Reproduction Via Genetics
- Breed Complementarity
- Heterosis
Breeding Systems
What is Crossbreeding?
Jim Gosey, Animal Science Department, University of Nebraska - Lincoln
Proceedings, The Range Beef Cow Symposium XIX December 6, 7 and 8, 2005, Rapid City, South Dakota
INTRODUCTION
Most ranchers know crossbreeding can increase output, but perhaps, don’t appreciate the potential 25% crossbred advantage in lifetime productivity of crossbred cows. Yes, you read that correctly; 25% crossbred advantage in lifetime productivity. In recent years many commercial cow herds have changed dramatically as producers have opted to repeatedly top-cross Angus bulls on their commercial cows resulting in loss of heterosis and loss of complementary breed effects. Some of the reasons for this shift are; 1) a desire to simplify breeding programs (perception that crossbreeding systems are too complex), 2) use of black hide color as a proxy for market quality, 3) the belief that high percentage purebred commercial cattle produce more uniformity and consistency, 4) effective marketing of the Angus EPD’s and carcass database, and 5) the Angus brand (CAB) impact (desire to get away from marketing commodity products).
A number of textbook crossbreeding systems are not “rancher friendly” in terms of management ease even though they deliver maximum heterosis. Additionally, dealing with grazing rotations, labor constraints and variable market targets require tough decisions that may tilt the crossbreeding system away from the original plan. Utilization of heterosis and breed differences in a crossbreeding system must be coupled with common sense ranch management in such a way that optimum (not maximum) heterosis is produced. There are some simplified crossbreeding systems that can meet this need very well.
Ranchers would be wise to crossbreed even if heterosis was zero, due to the complementary effects of matching strengths of one breed to offset the weaknesses of another breed. The opportunity to mate bulls and cows of different breeds or paternal/maternal lines to take advantage of complementarity is an important part of the total crossbred advantage. Just think back 40 years to what the Angus x Hereford cow did to match up the strengths of those two breeds and mask some of the weaknesses of each; that was complementarity!
The formation of composite breed types based on a multi-breed foundation is an attractive alternative to traditional crossbreeding systems. Composite breed types are based on matings among crossbreds of two or more breeds. Once a composite is formed, it can be managed as a straightbred in a one-pasture system with none of the problems associated with small herd size or fluctuation in breed composition.
DOMINANCE GENE ACTION PRODUCES HETEROSIS
Heterosis (hybrid vigor) is measured as the performance advantage of crossbreds over the average of their straightbred parents. Occasionally, crossbreds will perform better than either parental breed, however, heterosis should be measured against the average of the parental breeds. Heterosis can impact many traits, but is especially useful in improving performance in lowly heritable traits, such as, reproduction, early growth and fitness or lifetime productivity as shown in Table 1.
Table 1. Average Heterosis in Beef Cattle Traits
Trait % Heterosis Calf Crop Weaned 8 Wean Wt 13 Yearling Wt 4 Carcass Traits 3 Lifetime Productivity 25
On the other hand, highly heritable traits (above 40% heritability like some carcass traits) respond best to direct selection. Response to selection is due to additive gene action, thus the expression of a trait adds up in proportion to the number of beneficial genes. However, the variation in lowly heritable traits is accounted for mostly by dominance gene action and to a lesser extent by epistasis or gene interaction. The result of dominance gene action is the heterozygous gene pairs are superior to the homozygous gene pairs.
HETEROSIS = RECOVERED INBREEDING DEPRESSION
Maximum heterosis is realized in the first cross of distinctly different breeds. Subsequent backcrossing to either parental breed (such as in a rotational crossbreeding system) will reduce the expected amount of heterosis realized. Backcrossing to either parental breed will increase the level of inbreeding and thus reduce heterosis. Inbreeding (mating of related individuals, such as half-sibs) will “fix” more homozygous gene pairs and generally result in depression of production, particularly so in reproduction and fitness traits. Since all breeds are slightly inbred, the level of heterosis found in breed crosses is, in reality, due to the recovery of accumulated inbreeding depression.
The largest and most dramatic expression of heterosis is found in crosses between bos indicus (Brahman) cattle and bos taurus (European origin) cattle because they do not share any recent common ancestors.
Much effort has been devoted to research on developing inbred lines within a breed for the specific purpose of crossing them to generate line-cross heterosis. This research has failed to produce any useful heterosis between inbred lines within a breed other than to barely offset the initial losses due to inbreeding depression.
THE POWER OF MATERNAL HETEROSIS
Heterosis can be partitioned into three components; 1) individual heterosis, that found in crossbred calves, 2) maternal heterosis, that found in crossbred cows, and 3) paternal heterosis, that found in crossbred sires. By far, the most important of these is maternal heterosis, accounting for about 2/3 of the total crossbreeding advantage. Maternal heterosis has more impact because of the effect on reproductive performance through earlier puberty, higher conception rate, faster breed back, greater longevity and the maternal impact on calf performance. Individual heterosis generally accounts for the other 1/3 of the potential 25% increase in lifetime productivity and is realized due to early vigor resulting in more live calves plus greater early calf growth rate. Paternal heterosis does exist in mating ability but is rarely measured unless crossbred bulls are exposed to high numbers of cows (40 cows or more) in the breeding pasture. If bulls are only exposed to 25 cows and they are all pregnant, crossbred bulls have no opportunity to demonstrate their advantage in mating ability beyond changing the calving distribution. Most ranchers would consider stretching their bull power in this manner as an unnecessary risk, thus paternal heterosis is rarely measured.
CROSSBREEDING SYSTEMS
Table 2. Shows the heterosis produced by a two-breed rotational crossbreeding system, a three-breed rotational crossbreeding, and a rotational terminal system using a third or fourth unrelated breed as the terminal. The total amount of the crossbred advantage (combination of heterosis and complementarity) is shown in Table 3.
Table 2. Heterosis in Traditional Crossbreeding Systems
| Crossbreeding System | % Heterosis | |
| 2-Breed Rotation | 67 | |
| 3-Breed Rotation | 87 | |
| Rotation Terminal | 2X=67+100 | |
| 3X=87+100 |
Table 3. Crossbred Advantage in Traditional Crossbreeding Systems
| Crossbreeding System | % Crossbred Advantage | |
| 2-Breed Rotation | 16 | |
| 3-Breed Rotation | 20 | |
| Rotation Terminal | 24 | |
Additional breeds could be added to increase heterosis, but there is a realistic limit to the number of breeds that can be used since the management complications multiply as the number of breeds increase. For example, rotational crossbreeding systems require the breeds used to be similar in major traits areas, such as mature size, calving ease, milk production, etc. The number of breeding pastures needed increase in proportion to the number of breeds used in the system. The sire breed identity of each replacement heifer is needed in order to mate those heifers to bulls of a different breed, thus avoiding backcrossing and optimizing heterosis.
One of the major drawbacks of rotational crossbreeding systems is the substantial swing in breed composition that occurs between generations and also between years. Since two or more breeds of purebred bulls are used within a year, the resulting variation in breed composition is the primary reason that crossbreeding is perceived to result in more variation than straight-breeding programs. Table 4 shows a three-breed rotation program and the resulting breed composition for the three breeds. The average % breed composition hides the fact that there is large variation in breed composition from generation to generation, thus making it extremely difficult to assemble load lots of calves that are uniform.
Table 4. Breed Composition of a Three-Breed Rotation
% Breed Composition
| Generation | Breed of Sire | Breed A | Breed B | Breed C |
| 1 | A | 50 | 50 | |
| 2 | B | 25 | 50 | 25 |
| 3 | C | 12 | 25 | 62 |
| 4 | A | 56 | 12 | 31 |
| 5 | B | 28 | 56 | 16 |
| 6 | C | 14 | 28 | 58 |
| Average % | 31 | 29 | 40 |
Rotational-terminal crossbreeding systems are extremely effective in gleaning heterosis from a two or three-breed rotation to produce replacements and young crossbred females that are mated to terminal sires once they reach 5 or 6 years of age. Such a system harvests heterosis and the important other half of the crossbreeding advantage, namely complementary of breed differences. One of the drawbacks of rotational-terminal systems is they don’t fit small herds of cows. A three- or four-bull herd (90 to 120 cows) would be the minimum number needed to make a rotational-terminal system work. Obviously, if artificial insemination was used, some of the management and herd size considerations could be eased.
BREED EFFECTS ARE LARGE
A brief review of breed differences and biological types based on Germ Plasm Evaluation research at the Meat Animal Research Center (MARC) clearly shows within-breed, as well as between-breed differences, are large and that there is much overlap of trait distributions between breeds. However, it is also clear that breed means are truly different and the success (or failure) of crossbreeding programs may be decided when the choice of breeds is made for the foundation.
Table 5. Breed Group Efficiency of Gain To Different Endpoints (grams/mcal me)
| Breed | Time | Carcass Wt. | Retail Product Wt. | Marbling |
| Red Poll | 35 | 48 | 28 | 51 |
| Angus | 35 | 49 | 26 | 54 |
| Limousin | 47 | 54 | 57 | 47 |
| Gelbvieh | 40 | 49 | 49 | 45 |
| Simmental | 38 | 52 | 46 | 49 |
| Charolais | 40 | 53 | 50 | 49 |
| MARC 1 | 39 | 51 | 45 | 48 |
| MARC 2 | 37 | 52 | 37 | 52 |
| MARC 3 | 35 | 50 | 30 | 53 |
Time=207 d, Carcass Wt. =734#, Retail Product Wt = 463#, Marbling = 4.0small
Table 5. Points out the opportunity for breed complementarity in efficiency of British and Continental breeds when fed to either a time, carcass weight, retail product weight or marbling constant slaughter endpoint. Note the change in breed ranking for efficiency of gain at the different endpoints. British breeds are more efficient when fed to a marbling constant endpoint and Continental breeds are more efficient when fed to a time, carcass weight or retail product constant endpoint. The MARC II composite (1/4 each Angus:Hereford: Simmental:Gelbvieh) provides the best complementary fit for efficiency of gain to both a marbling and carcass weight constant endpoint.
Also, research at MARC on efficiency of feed use in nine purebred breeds of cows indicates breeds that excel at low levels of dry matter feed intake (generally the British breed types) lack the productivity (growth and milk production) to excel at high dry matter feed intake. Likewise, highly productive breeds (generally the Continental breed types) are the least efficient when limited to low levels of dry matter feed intake. Thus, fitting these major breed differences to the carcass targets for progeny and to the feed environment for cows is critical to the success of crossbreeding programs.
CROSSBREEDING WITH COMPOSITES
While hybrids and composites are both crossbreds, hybrids are generally considered to be F1 or first crosses of purebred parents and composites are the result of matings among crossbred parents. The composite seedstock breeder must take special care to plan the formation of the composite to avoid inbreeding, thus a “closed composite” requires a large herd size, estimated at 25 sires per generation to hold inbreeding to less than .5% per generation. A composite seedstock breeder that uses an “open composite” approach has a much lower requirement for herd size since new sires (and perhaps breeds) are continually being evaluated and introduced, probably via AI, thus holding the inbreeding level to a minimum. Existing breeds of cattle are mildly inbred lines and to the extent that heterosis is due to dominance gene effects, heterosis is the recovery of accumulated inbreeding depression, thus managing inbreeding in composite breed formation is critical to success.
Table 6. Example Crossbreeding Systems
| Crossbreeding System | Minimum % Breed A | Maximum % Breed B | Percent F1 Hybrid Vigor |
| Rotate Purebred A & B bulls | 33 | 67 | 67 |
| Rotate Purebred A, B & C bulls | 14 | 57 | 86 |
| Rotate F1 AxB and F1 CxD bulls | 17 | 33 | 83 |
| Composite AxBxCxD bulls | 25 | 25 | 75 |
| Composite AxB bulls | 50 | 50 | 50 |
| Composite A x (BxC) bulls | 50 | 50 | 63 |
| Rotate F1 AxB and F1 AxC bulls | 50 | 50 | 67 |
Composite breed types do not sustain as high of level of heterosis as do the traditional rotation crossbreeding systems as seen in Table 6, however, composites do allow for more complementarity between breeds. Several examples are shown in Table 6 that level the contribution of a given breed (Breed A in this example) or several breeds. Table 7 demonstrates the impact of the number of breeds and the impact of equal contribution of each breed to the foundation generation. The number of breeds used in the foundation of a composite accounts for most of the heterosis retained, however, the heterosis is reduced as the contribution of each breed to the foundation is less than equal. Heterosis retained is proportional to the heterozygosity retained in a cross and is equivalent to (n-1/n), where n = the number of breeds. So a four-breed composite would produce 75% heterosis and that level would be maintained over time. The initial loss of heterosis is due to loss of heterozygosity which occurs between the F1 and F2 generations but is maintained in subsequent generations of crosses in a composite.
Table 7. Composite Heterosis By Mating Type
| Number of Breeds | Breed Foundation | % Heterosis | % Crossbred Advantage |
| 2 | 1/2:1/2 | 50 | 12 |
| 5/8:3/8 | 47 | 11 | |
| 3/4:1/4 | 38 | 9 | |
| 3 | 1/2:1/4:1/4 | 63 | 15 |
| 3/8:3/8:1/4 | 66 | 15 | |
| 4 | 1/4:1/4:1/4:1/4 | 75 | 17 |
Some breeders have assumed that variation in composite populations is greater than that found in purebred populations, however, in a definitive study of the three composite lines at MARC and their parental purebreds, there was no significant difference in the coefficient of variation for reproduction, production or carcass traits measured (Table 8).
Table 8. Coefficients of Variation For Purebred vs. Composite Steers
| Trait | Purebreds | Composites |
| Birth Wt. | .12 | .13 |
| Wean Wt | .10 | .11 |
| Carc. Wt. | .08 | .09 |
| Retail Product % | .04 | .06 |
| Marbling | .27 | .29 |
| Shear Force | .22 | .21 |
Another criticism that has been leveled against composites is they lack the accuracy of EPD’s found in many purebred breed evaluations. This points out the need for multiple breed evaluation to be expanded between the most widely used breeds in commercial production.
Recently (Fall 2005) the Gelbvieh, Brangus, Limousin, Salers and Red Angus associations have created a new company, Performance Registry Services. The goal of this joint venture is to provide commercial producers with National Cattle Evaluations by delivering a single suite of EPD’s for all the participating breeds on a single base. This joint venture of breed associations will provide Total Herd Reporting of all cattle regardless of breed combination and data processing for all their members while allowing for individual breeds to maintain their own identity. Along with the multi-breed EPD’s there will be decision support software, search engines and a centralized data warehouse for commercial producers to use these tools. The result of this effort will be to make it easier for producers to evaluate breed inputs into crossbreeding programs.
The commercial user of composite breed types has to worry about few of the constraints that the composite seedstock breeder encounters, as they can be managed as a straightbred in a one-pasture system. Composite breeds offer the opportunity to use genetic differences among breeds to achieve and maintain the performance level for such traits as climatic adaptability, growth rate and mature size, carcass composition, milk production, and fertility that is optimum for a wide range of production environments and market scenarios. Further, composite breeds may provide herds of any size an opportunity to use heterosis and breed differences simultaneously.
Composites offer an opportunity to counter the antagonism between USDA Quality Grade and Yield Grade as shown in Table 9. The often-stated goal of the beef industry is to produce finished cattle that are at least 70% USDA Choice or better, 70% Yield Grade 1 & 2 and have zero defects or zero “out” cattle. This 70-70-0 target is difficult to achieve with either British or Continental breeds alone, however, a blend of these two types as found in the MARC II (1/2 Continental:1/2 British) does a much more acceptable job of meeting the 7070-0 target. Thus a composite can actually lower the risk of non-compliance to a market target.
Table 9. Conformance of Breed Types to Carcass Targets
| Item | British | Continental | Marc I | Marc II | Marc III |
| %Y 1&2 | 38 | 89 | 83 | 56 | 53 |
| % CH - | 70 | 30 | 43 | 55 | 66 |
| % Non-Conformance To 70 – 70 – 0 Target | |||||
| Yield Grade | 32 | 14 | 17 | ||
| Quality Grade | 40 | 27 | 29 | 21 | |
| Total | 32 | 40 | 27 | 29 | 21 |
Careful selection of foundation sires used in the development of a composite can further move a herd toward meeting market targets. Table 10 shows six lots of steers born at the Gudmundsen Sandhills Laboratory near Whitman, Ne. which were sired by bulls produced in the University of Nebraska Teaching herd. Unlike the MARC Germ Plasm Utilization project where bulls were sampled across a broad spectrum of each breed, the foundation sires in the UNL Teaching herd were selected using EPD’s to be above average in calving ease, average in milk production, average or below in mature size, and above average in marbling and other carcass traits. The result is steers on average that are 87% USDA Choice or better and 66% Yield Grade 1 & 2. Several of the individual lots of cattle quite easily surpassed the 70-70-0 market target.
Table 10. Calves sired by University of Nebraska Composite bulls.
| Date | # | Wt | Fat | REA | YG | %Y1:2 | %Ch |
| 6/05 | 37 | 836 | .54 | 13.2 | 3.19 | 49 | 97 |
| 5/05 | 45 | 823 | .57 | 13.8 | 3.02 | 49 | 84 |
| 0/05 | 89 | 795 | .51 | 13.5 | 2.83 | 62 | 85 |
| 3/05 | 22 | 802 | .41 | 14.6 | 2.34 | 82 | 91 |
| 3/05 | 24 | 729 | .49 | 13.0 | 2.74 | 75 | 96 |
| 12/4 | 53 | 809 | .40 | 14.5 | 2.35 | 89 | 81 |
| AV. | 270 | 802 | .49 | 13.8 | 2.77 | 66 | 87 |
SUMMARY
Heterosis and complementarity are powerful forces that combine to produce the total crossbred advantage of beef cattle crossbreeding. This crossbred advantage can amount to as much as 25% greater lifetime productivity (pounds of calf weaned per cow exposed) for crossbred cows as compared to straightbred cows. Some commercial cowherds have drifted towards straightbred Angus herds in an attempt to achieve management simplicity, greater uniformity in their cattle, and to pursue a premium (non-commodity) product. The result of this shift is the loss of most of the heterosis that once existed in many of our commercial
cowherds. Loss of heterosis shows up in the same lowly heritable traits that would be associated with inbreeding depression, namely reproductive, fitness and longevity traits. Thus, the price paid for loss of heterosis occurs as a number of very small losses that when added up can amount to a substantial sacrifice in lifetime productivity (25%).
Traditional crossbreeding systems (rotations & rotation-terminals) are very efficient in maximizing heterosis but are more complex than many producers would like. Perhaps the availability of estrus synchronization protocols for timed AI will assist some commercial producers in using some of the traditional crossbreeding programs in the future. One-pasture crossbreeding programs exist that can deliver adequate (not-maximum) heterosis, are simple to manage, utilize breed differences (complementarity), can be designed to produce uniform calf crops and can help avoid several important genetic antagonisms. Composite breeds must be carefully formed with the same attention to breed choices and sire selection that is used in straight breeding programs. However, once formed the commercial user of composites can manage a composite crossbreeding program with greater management ease than traditional crossbreeding systems. One-pasture crossbreeding programs offer commercial producers a practical tool to enhance management effectiveness and increase profitability.
Improving Cow Herd Reproduction Via Genetics
By Wade Shafer, PhD., ASA Executive Vice President
Editor’s Note: This article was originally published in March 2008 issue of the SimTalk written by Wade Shafer, PhD., Drs. Lauren Hyde and Jackie Atkins provided updates for reprint.
A beef cow’s job is not an easy one. She is expected to conceive at slightly over one year of age, to calve by the time she is two, and rebreed shortly after that while weaning a healthy, viable calf. Furthermore, we demand that she consistently repeats this cycle for the rest of her life — one stumble and, hasta la vista, baby!
To be sure, producers are best served when the cow successfully performs her task for many years, as the longer her productive life, the more profitable she is to the enterprise. Is there anything that can be done to help her out? Certainly, there are environmental factors we can manage that will give her a leg up. For example, by providing adequate nutrition and a proper vaccination regimen and mating her to easy-calving sires (particularly when she is young), we increase the odds of her success. While a cow’s environment has a substantial impact on her reproductive performance, her genetic makeup can too. This article explores the genetics of female reproduction and offers suggestions on how to improve the reproductive performance of your cow herd via genetics.
Crossbreeding
The obvious place to start a discussion about the genetics of female reproduction is the factor that far and away has the greatest effect on it — crossbreeding. It has long been recognized that crossbreeding enhances virtually all aspects of reproductive performance. Studies too numerous to list here have established the reproductive superiority of crossbred over straightbred cows.
In one of an abundance of studies with similar findings, scientists at the Meat Animal Research Center (MARC) concluded that two-breed rotational cross cows produced 20% more calves over their lifetime than straightbreds due to the favorable impact of heterosis on dam fertility/longevity and calf survivability brought about by the improved calving and mothering ability of the dam (Cundiff et al., 1992). Furthermore, they estimated that when mated to a bull of another breed, the two-breed cross cows would wean 36% more weight over their lifespan than straightbred cows raising straightbred calves. The dramatic increase is attributable to the positive influence of heterosis on reproduction and production in the dam and well as increased growth and survivability in their calves.
Given the overwhelming evidence of the crossbred cow’s reproductive supremacy and the fact that reproduction is a major piece of the profitability puzzle (by most accounts exceeding all other functions by a wide margin of relative importance), it is difficult to conceive of a situation where a commercial enterprise would not benefit financially from a crossbred cow herd.
Are we implying that selecting animals within a breed for reproductive performance is not a worthwhile endeavor? No! Reproductive progress can be made via selection (which we will address later); however, it would take years of intense selection within a breed to yield the kind of improvement that can be achieved in one fell swoop by simply crossbreeding.
Therefore, crossbreeding makes a logical cornerstone in any effort to enhance cow herd reproductive performance. With crossbreeding as the foundation, the selection of superior animals of multiple breeds as inputs to the crossbreeding system can be considered a supplemental means of further boosting reproductive function; however, identifying reproductively superior animals has its challenges, as we will explain.
Indirect Selection
Because the assessment of a cow’s reproductive performance is generally determined later in her life, it seems logical to look for early indicators to hasten the process. For example, it is a commonly held belief that females with a propensity toward fatness will excel reproductively.
Though research has shown that increased fatness, to a point, is strongly and favorably associated with reproductive performance on a phenotypic scale, the few attempts to assess the relationship on a genetic level shows an unfavorable, though weak, relationship. Using data from the Red Angus Association of America (RAAA), researchers at Colorado State University (CSU; Beckman et al., 2006) derived genetic correlations ranging from -.12 to -.22 between body condition at various ages and Stayability (by industry convention, the probability of a cow remaining in the herd through 6 years of age). At the American Simmental Association (ASA), we have found a correlation of -.19 between an animal’s genetic propensity for backfat in the feedlot and their inherent Stayability. We (ASA) have also calculated a -.11 genetic correlation between backfat and heifer pregnancy (the likelihood of a heifer being pregnant at the end of the breeding season) using RAAA data.
Admittedly, these unfavorable correlations between fatness and reproduction may seem illogical. We have all seen a higher proportion of thin cows open at pregnancy test time. Keep in mind, however, that the aforementioned correlations are genetic correlations. The relationships we actually observe, i.e., phenotypic correlations, are influenced by a combination of underlying environmental and genetic relationships. There is little question that females within a herd lucky enough to experience an environment for increased body condition (e.g., extra energy intake) are likely to have better reproductive performance than their herd mates. Furthermore, this strong and positive environmental relationship between fat and reproduction apparently overwhelms what appears to be a slightly negative genetic relationship — yielding the strong, favorable phenotypic relationship we typically observe.
Frankly, there is not enough evidence about the genetic relationship between fatness and reproductive function to make recommendations based on it at this time; however, though it may fly in the face of conventional wisdom, it appears that selecting “easy-fleshing” genotypes will not gain us ground reproductively.
Scrotal circumference has been considered as a predictor of female reproductive performance. Though the preponderance of evidence indicates a strong to a moderately favorable relationship between scrotal circumference and age at puberty in related females, research is less clear on the relationship between scrotal circumference and subsequent measures of reproduction. In a study based on a large population involving several breeds at the MARC, Martinez-Velazquez et al. (2003) found a slightly unfavorable (.15) relationship between scrotal circumference and age at first calving and no relationship between scrotal circumference and first pregnancy, first calving, and first weaning rates. Their conclusion was that selection on scrotal circumference would not be effective in improving female reproduction. These findings are in agreement with some studies and contradicted by others. For those interested, Martinez-Velazquez et al. (2003) provides an excellent literature review on the subject. Given the conflicting evidence, it may not be advisable to base selection decisions on scrotal circumference with the intent of enhancing maternal reproduction.
As for other traits that may be related to reproductive function, Rogers et al. (2004) found that increased levels of milk EPD increased the risk of females being culled. This finding is consistent with ASA data showing an unfavorable (-.15) genetic correlation between milk and Stayability. Other ASA genetic correlations of note are -.26, .40, and -.19 between Stayability and mature weight, maternal calving ease and marbling, respectively. Based on these findings, we would expect females that are inherently lower milking, smaller at maturity, easier calving, and less marbled to stay in the herd longer; however, none of these relationships is strong enough to make a sizable impact on Stayability by selecting for them. Furthermore, other than mature weight, because of its strong relationship to early growth, determining the genetic level of a young heifer for these traits by simply observing them (which is what most commercial producers are limited to) is not possible. Therefore, a different tactic will be required if we wish to improve reproductive performance via selection. Namely, select for it directly — which, as we will point out, is not a trivial task.
Direct Selection
A well-entrenched view of both commercial and seedstock producers is that the “cows left standing” after culling on the components of reproduction (e.g., pregnancy status and calf loss) are genetically superior. By extension, it is presumed that a great deal of progress in reproduction is made through rigorous culling and the retention of heifers out of dams making it to advanced ages. Though this may seem like a reasonable eduction, it is generally not the case.
Unfortunately, little genetic headway is made by simply culling cows that do not achieve reproductive thresholds. This may seem counterintuitive. Why wouldn’t getting rid of the offenders improve your genetics for reproduction? The main reason lies in the fact that measures of reproduction tend to be lowly heritable (estimates typically run between 5-20%). And, with lowly heritable traits, an animal’s own performance is not a good indicator of its genetic level for the trait. Therefore, many open culls may be genetically above average or even superior for reproduction. By the same token, several cows kept because they are bred may be genetically inferior for it — certainly not an outcome that will yield much improvement.
So, how do we directly select for reproduction? Because a cow’s reproductive performance is expressed later in life, and even then it only provides a very cloudy picture of her genetic merit, are we relegated to making little to no selection progress for reproduction? Heck, no! We can clear the clouds with reproductive EPD.
Though EPD always provide the best estimate of an animal’s genetic merit, they are especially valuable when applied to low-heritability traits. This is because, when an animal’s own record is a poor indicator of its genetic makeup, gathering information on its relatives is the only means we currently have of getting a clear picture of the animal.
You may ask yourself, “If an animal’s own performance does not tell us much, what can be gained by records on its relatives?” It is not that a single relative record brings much to the mix (obviously it adds even less than the animal’s own record); it is that there is strength in numbers — an animal can have many relatives with records, but only one record on itself. Through the use of EPD, we utilize information on all of an animal’s relatives and, in doing so, chip away at the cloud with each record that flows in.
With a low-heritability trait expressed later in life, like reproductive function, the cloud clears slowly — but it will clear. In fact, if an animal has enough progeny records, we can see its genetic merit for reproduction as clear as a bell.
Fortunately, the seedstock industry now has EPD that are, for the most part, direct measures of reproductive function: Stayability (STAY) and heifer pregnancy (HP). Researchers at CSU developed STAY (Snelling et al., 1995) and HP (Doyle et al., 2000) EPD, and the RAAA implemented them into the association’s national cattle evaluation a few years later. Since its development, STAY has undergone several revisions. Most recently, the ASA released the industry’s first multi-breed STAY evaluation, which incorporated genomic data in a single-step random regression model.
Though STAY and HP have potential shortcomings (e.g., seedstock breeders’ culling practices are probably not in step with the commercial industry’s, and breed association culling records tend to be sketchy), they are the most effective selection tools available for improving reproductive function. What’s more, based on computer simulation efforts by retired USDA scientist M.D. MacNeil, the economic impact of Stayability when selecting a sire for female replacement is nearly twice that of the next closest trait, while the relative importance of heifer pregnancy is on par with the most important carcass or growth traits (personal communication) — so these reproductive EPDs certainly warrant a great deal of attention in the selection process.
Most commercial producers do not have the luxury of using STAY or HP EPD to select replacement females; however, if you select sires with superior EPD in these areas, the reproductive function of your cow herd is likely to improve over time. Given their relationship to Stayability, you may also gain some reproductive ground by selecting sires with lower milk, smaller mature size and better maternal calving ease EPD. Another option to consider for commercial producers is the commercial option of the American Simmental’s Total Herd Enrollment. The commercial option predicts EPD on commercial females and coupled with the Cow Herd DNA Roundup provides genomically enhanced EPD to commercial females.
Summary
In closing, we must reiterate that crossbreeding needs to be at the center of any effort to improve the reproductive function of your cow herd. The dramatic impact of heterosis on reproductive performance is crystal clear — no herd should be without it! Though reproductive improvement through selection is possible, it is generally limited to utilizing reproductive EPD when selecting your herd sires. By combining crossbreeding with the selection of superior sires you will position your enterprise to excel in the most vital area of beef cattle production — cow herd reproduction.
Literature Cited
Beckman, D. W., S. E. Speidel, B. W. Brigham, D. J. Garrick, and R. M. Enns. 2006. Genetic parameters for stayability and body condition score in beef females.
Proc. West. Sect. Am. Soc. An. Sci. 57:93-95. Cundiff, L. V., Nuiiez-Dorniguez, R., Dickerson, G. E., Gregory, K. E., and R.
M. Koch. 1992. Heterosis for lifetime production in Hereford, Angus, Shorthorn, and crossbred cows. Journal of Animal Science. 70:2397-2410.
Doyle, S. P., Golden, B. L., Green, R. D., and J. S. Brinks. 2000. Additive genetic parameter estimates for heifer pregnancy and subsequent reproduction in Angus females. Journal of Animal Science. 78:2091-2098.
Martinez-Velazquez G., K. E. Gregory, G. L. Bennett and L. D.
Van Vleck. 2003. Genetic relationships between scrotal circumference and female reproductive traits. Journal of Animal Science. 81:395-401.
Rogers, P. L., Gaskins, C. T., Johnson, K. A., and M. D. MacNeil. 2004. Evaluating longevity of composite beef females using survival analysis techniques. Journal of Animal Science.
82:860-866.
Snelling, W. M, Golden, B. L., and R. M. Bourdon. 1995. Within-herd genetic analyses of stayability of beef females.
Journal of Animal Science. 73:993-1001.
Breed Complimentarity
Breed Complementarity
Heterosis
Hybrid Seedstock
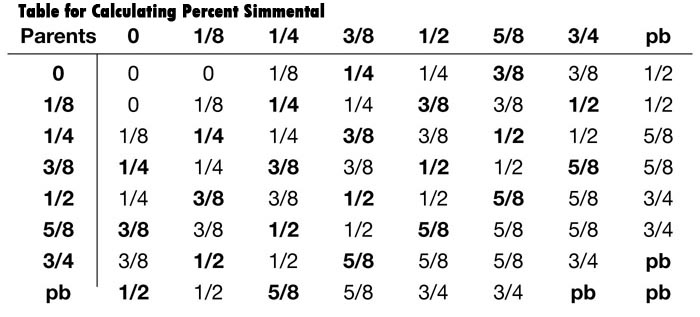
Table for Calculating Percent Simmental
Using Composite Seedstock
The use of composite seedstock in beef cattle breeding operations is growing at a fast rate. With the introduction and advancement of the IGS Multibreed Genetic Evaluation, in addition to an understanding of the benefits of composite seedstock, hybrid bulls and females are popular items in today’s commercial world.
Hard hitting things-it is easy, it has exploded in popularity, its in the incorporation of the genes of econ relevance, percent heterosis and not a lot of breeding pastures.
By Rick Bourdon Colorado State University
There is a growing interest among commercial cattle producers in the use of composite cattle as an alternative to crossbreeding. Naturally, there is a parallel interest among seedstock breeders, many of whom are currently purebred breeders, in producing composite seedstock. What these breeders need to understand, however, is that there are fundamental differences between breeding composite seedstock and breeding purebreds. My purpose in this set of articles is to make these differences clear.
There are two underlying reasons why composite and purebred breeding differ. The first is that in composite breeding there are two distinct breeding stages: (1) forming the composite, and (2) breeding the composite once it is formed. The second reason is that in composite breeding we are interested not only in improving breeding value through selection¾ our sole genetic goal in purebred breeding¾ but also in maintaining a high level of hybrid vigor. These basic, theoretical differences between the two types of breeding spawn a number of practical differences. In this first article, I look at these differences in the context of composite breed formation. In the sequel, I look at them in the context of breeding the formed composite.
If you are not interested in creating a new composite breed, but simply want to breed cattle of an existing composite breed, the following discussion of composite breed formation will be of little value to you. We are at a point in time, however, when many people are contemplating creating new composites. Some of these breeds are in the initial stages of breed formation. Others are just a twinkle in a breeder's eye. In either case, the following points should be helpful.
Improving breeding value through between-breed selection
Selection of the breeds and the proportions of those breeds going into a composite is the critical step in composite breed formation and may well determine whether a breed succeeds or fails. If composite breeders do a carefully researched and conscientious job of between-breed selection, then the newly formed composite will not need much genetic change¾ change that can now only be achieved through slow-paced within-breed selection. In other words, if the composite is put together in such a way that it exhibits close to optimum performance in the economically important traits when it is first formed, then any genetic change following breed formation can be considered fine tuning. Following are some ideas to keep in mind when designing a composite breed.
Define how the breed will be used. When companies come out with a new product, they almost always know exactly how the product will be used. Without that knowledge, it would be hard to both design and market the product. The same is true with composites. We need to know how the composite will be used by commercial producers. We need to define the geographical areas and environment/management niches appropriate for the composite breed.
It is also important to define how the breed will be used in the context of mating systems. Most composites are likely to be "all-around" or general purpose breeds because that is the kind of animal
required for the simplest or "pure" form of commercial composite breeding systems. These breeds should be strong in all traits, or at least not weak in any important trait. They must combine good maternal characteristics with carcass quality and cutability. Other composites may be specialized maternal breeds designed to be bred to a terminal sire. They will probably be smaller, strong in maternal traits and adaptability, and have carcass characteristics¾ typically carcass quality attributes¾ that will complement the carcass characteristics of the terminal sires. We may see specialized paternal composite breeds¾ terminal sire breeds for producing either the ultimate market animal or simply live calves out of first-calf heifers. Whatever the potential use for a composite breed, those who form the breed should keep that use in mind when designing the composite.
Aim for optimums. There may not be optimum levels of performance for all traits, but common sense tells us that there are optimum levels of performance for some traits. Milk production and mature size come to mind right away. Clearly, there are limits to the amount of milk a beef cow can efficiently produce and limits to the mature weight she can efficiently maintain. Admittedly, optimum levels for traits like these are not easy to determine, but to the extent that they can be defined, we should try to come close to them in composite breed formation. In other words, choose breeds and breed proportions in such a way that optimal levels of performance are reached in the first composite generation.
Exploit breed differences. One of the unique aspects of composite breeds is that they can incorporate component breeds that most commercial producers would be reluctant to use in any kind of a crossbreeding system involving purebred sires. Consider the Jersey breed. Jerseys have a lot to offer: excellent fertility and milk production, lovely udders, great paternal and maternal calving ease, low absolute maintenance costs, and highly marbled, tender meat. They also have a downside: slow growth rate, poor feed efficiency and carcass cutability, small carcasses, and a nasty disposition (in bulls). The negatives are enough to keep most commercial producers from using Jersey sires. However, a composite containing the right dose of Jersey combined with other component breeds that compensate for the Jersey's weak points could be really useful. In effect, by "mixing and matching" breeds in composite formation, it is possible to make use of complementarity¾ the production of a more desirable offspring from the mating of parents that are genetically different from each other but have complementary attributes.
One of the most persistent misconceptions about composites is that they lack uniformity. Producers assume that if hybrids are mated to hybrids, the offspring will vary across the board. We know from theory and from experimental results that this is not true. Composites are as uniform as purebreds for almost all economically important traits. Only in those traits affected by few genes¾ coat color is the best example¾ will composites be clearly less uniform. Color uniformity may be something to consider when choosing component breeds. This may seem silly¾ we don't eat the coat, after all, but it is amazing how uniformity of color affects people's perception of uniformity in general.
Improving breeding value through within-breed selection
Choose the best foundation animals. The choice of what breeds to include may be the single most critical step in composite breed formation, but the breeder's responsibility does not end there. We all know there can be tremendous variability within breeds. It is important, therefore, that composite breeders be selective in their choices of foundation animals. It would not do, for example, to pick just any Hereford or Jersey bulls simply because they were purebred and available. What if they were not representative of those breeds or had some glaring faults? Furthermore, we sometimes want to include only certain types within a component breed. I can think of several breeds that have a lot to offer, but are (in my opinion) too big at birth and maturity. However, there are bulls within those breeds with atypically low birth and mature weight breeding values, and these are the bulls I should hunt for. If color uniformity is a concern, foundation animals should be screened for their color genotype as well. A good job of composite breed formation means a good job of both between- and within-breed selection.
Maintaining hybrid vigor
Sample widely. The key to maintaining hybrid vigor in a composite is to avoid inbreeding. The way to prevent inbreeding at the breed formation stage is to establish as broad a genetic base as possible. From a practical standpoint this means including in the foundation population a number of unrelated sires or daughters of unrelated sires from each component breed. To see why, consider the extreme case of a four-breed composite in which one component breed's contribution came in the form of semen from a single sire. Every member of the first generation of composite animals will be a grandson or granddaughter of that bull. He will appear repeatedly in the pedigrees of future generations, and the result is an increase in inbreeding.
What is the minimum number of sires needed to represent a component breed? I don't really know. One is too few. Four or five, if they are truly unrelated, should probably be adequate. A dozen would be plenty.
Other concerns
Order of crossing. Say you were designing a four-breed A% B% C% D composite. Would the order of the initial crosses make a difference? In other words, would it matter whether the first composite generation was (A% B)% (C% D), (A% C)% (B% D), or (A% D)% (B% C)? The answer is yes and no. If the component breeds differ in maternal ability and(or) if specific two-breed crosses differ in hybrid vigor, the performance of the first generation of composites and, to a lesser degree, the second generation will be affected by the order of crossing. The performance of the third and more advanced generations should be unaffected, however.
Say you were designing a four-breed A B C D composite. Would the order of the initial crosses make a difference? In other words, would it matter whether the first composite generation was (A B) (C D), (A C) (B D), or (A D) (B C)? The answer is yes and no. If the component breeds differ in maternal ability and(or) if specific two-breed crosses differ in hybrid vigor, the performance of the first generation of composites and, to a lesser degree, the second generation will be affected by the order of crossing. The performance of the third and more advanced generations should be unaffected, however. Complexity. The biggest advantage of using composites is that they are easy to manage. Unfortunately, the same cannot be said for creating composites. In assembling a four-breed composite you could be dealing with purebreds of all four breeds, two two-breed crosses, and full composites¾ all at the same time. Creating a composite breed is not something to be entered into lightly.
Time required. Composite creation can be time-consuming too. I estimate that starting from scratch, it takes a decade to make the crosses for a four-breed composite and replace the foundation animals completely with composites. If you are getting on in years, composite development may be like planting shade trees¾ the next generation will be the first to enjoy the fruit of your labor. On the other hand, if you already have a set of two-breed cross females and can find the right two-breed F1 bulls, you can have four-breed composite calves on the ground next year.
In the next article in this series I discuss breeding composites once the composite breed has been put together.